So as I mentioned earlier, ( https://schollsvalley.com/notes-from-the-farm-21-september-2025-climate-adapted-willows/ ), willows are absolutely essential to ecosystem functions in the Willamette Valley, and are especially vital for healthy wetlands and streams. They are also taxonomically and ecologically complex. The selective forces that maintain the identities of various willow populations (“species”) are subtle, and barriers to hybridization among them are porous, leading to frequent F1 hybrids, back-crosses and likely many variously introgressed populations.
So what to do? Chill and enjoy it! Also, let’s be sure to appreciate what this state of affairs means for climate adaptation of willows, and maintenance of their critical ecological functions in a dynamic world.
If we take an open-minded approach to our Willamette Valley willows, they are actually a pretty tractable group. You are unlikely to encounter more than eight species in our little Valley, and a process of elimination using simple keys usually gets you to a reasonable taxonomic call for most specimens. For those that don’t fit in any box well, it is usually possible to at least guess at their parentage. For instance, F1’s and later-generation hybrids between Salix hookeriana and S. scouleriana appear to be quite common locally. Often, putatively hybrid individuals exhibit a number of features that are intermediate between these two species. Sometimes even the distinctive S. sitchensis appears to be involved in some intermediate specimens. So there is a lot going on, but careful observation can usually yield a sensible understanding.
I’m going to step back here and write briefly about hybrids in general, and my sense of them. Hybrids have always interested scientists, plant breeders and plant enthusiasts alike, for obvious reasons. They happen in plants with a frequency that exceeds that in animals. This likely has to do with the open system of growth and organization of plants and the closed, more complex system of development in animals; e.g. an extra branch on a tree may be inconsequential or even adaptively advantageous in some habitats, whereas an extra forelimb on a tetrapod is probably lethal. So plants have greater freedom to explore genetic and phenotypic spaces, which allows them to key into varied and dynamic niches as they appear in the landscape. This exploration is strongly promoted through the processes of hybridization and introgression (see for example Hamilton et al 2013 https://nph.onlinelibrary.wiley.com/doi/full/10.1111/nph.12055 ).
The functions of hybrids in the real world are central to my own research, and I have come to the conclusion that the occurrence of hybridization and introgression in plants is much more than just interesting. It has been gratifying to find common ground with other researchers on this point, and particularly, to come to unifying conclusions, using molecular as well as old-fashioned ecological and morphological study, with previous research. The fact that molecular evidence more often corroborates than refutes the work of 20th-century greats like Grant, Dobzhansky and Stebbins, is both powerful and empowering. The truth is that we can make good inferences using field observations, histology and well-conceived ecological experiments. We don’t always need to look to the molecules and computers to tell us what’s going on. Besides, if there is morphological and taxonomic confusion in a group, frequently the molecular picture is equally confounding. Don’t get me wrong – molecular science is great, and has certainly yielded many new insights in the life sciences, including in my own work. But it’s not the end-all-be-all answer to intractable taxonomic problems that we might have expected 40 years ago. And for field ecologists, that’s good news. A pair of eyes and a decent human brain remain your best tools to get you around in the woods.
Back to willows! We have been propagating willows from seed for nearly twenty years and have perfected the art of producing big crops of large, genetically diverse seedlings. These crops have contributed to the restoration of thousands of acres of local natural areas, including the Sandy River Delta, Clean Water Services riparian work in the Tualatin Basin and many other projects and restoration programs. But these plants remain underutilized. More on this in a future post.
One fantastic benefit we have in this large-scale propagation work is the opportunity to observe tens of thousands of individuals of known parentage, with multiple species and accessions side-by-side. This is always helpful to us in gaining a better understanding of these species – their comparative phenology, morphology, seed ecology, etc. And sometimes we learn something about inter-species interactions, as was the case this year with the unexpected emergence of an F1 population resulting from apparent interactions between Salix sitchensis and S. prolixa.
These two willows are morphologically very distinctive. Below is a table of just a few generalized leaf characters of the two parental species:
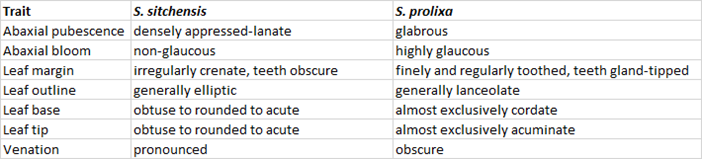
The putative F1’s appeared exclusively among a stand of Sitka willow, so S. prolixa is the presumed pollen parent in this case. The F1’s are taller than the average of their Sitka cohort, and much taller than the adjacent prolixa stand. But if it weren’t for the differences in pubescence, it would be challenging to be certain the F1’s weren’t just a seed cross-contamination issue in two adjacent blocks. Our prolixa is exclusively glabrous, and our Sitka is invariably densely woolly on abaxial leaf surfaces. The F1’s, although they appear more generally allied to S. prolixa, are nevertheless clearly intermediate in pubescence, with scattered appressed hairs and therefore distinctly different from either parent. In fact, close examination reveals that they are intermediate for all of the above leaf characters, and also intermediate for twig and coloration traits (not listed). Here is a photograph of leaves from ten individuals each of Sitka (top), the F1 hybrid (mid) and prolixa (bottom):

Pretty cool, huh? But also so much more. This is a beautiful and clear example of the first step in interspecific interaction. Where things go from here involves genetics (are the F1’s themselves fertile? How fertile are they? Are any later-generation hybrids stable?) and ecology (are the F1’s and later-generation hybrids – if any – competitive in any available habitat? Are there traits of either species that could be beneficial to the other that could introgress populations and even become fixed?). These ecological questions are all the more relevant in light of climate change and adaptation. Under a changing climate, most currently well-adapted populations are likely to become stressed and vulnerable to displacement by better-adapted ones. This opens the door to novel combinations. And nature has no more expedient means to generate novel combinations than hybridization.
We are going to segregate the putative F1’s and grow them in a block so we can observe this new population as it matures. We might expect some disruption of sex expression, or other evidence of reduced fertility. But given what we know, including evidence already of some level of heterosis, as well as the general promiscuity of willows, it is likely that at least some of these plants will be both competitive and fertile. Which gives them a chance to be relevant to the future trajectory of willow evolution here in the Willamette Valley.
And we have a unique chance to see if this might be so.
Until next time,
-George